Porchetta 2.0 – hidegfüstös, roppanós bőr protokoll
Feltekert, szaftos császárhús, hideg füsttel, fokhagyma pasztával és a ropogós bőr tudományos pontossággal kivitelezve karalábépüré ágyon (hidegfüstös karalábéval) rozmaringgal szuvidált, vajban pirított burgonyával
🕒 Elkészítési idő: 20 + 180 perc
👨👩👧👦 4 adag
🛒 Hozzávalók
Hús és fűszerezés:
1 kg császárhús (bőrrel)
8 gerezd fokhagyma
10 ág friss kakukkfű
2 ág rozmaring
1 csokor zsálya
6 g reszelt citromhéj
só, bors ízlés szerint
2 ek kacsazsír
néhány csepp citromlé
Zöldségalap:
240 g sárgarépa
240 g vöröshagyma
100 ml víz
100 ml száraz fehérbor
👩🍳 Elkészítés
- Előkészítés: A császárhúst hosszanti irányban kinyitom, a porcos részeket eltávolítom, és a bőrt sűrűn megszurkálom, hogy a zsír könnyen távozhasson.
- Fokhagymapaszta: A fokhagymát, friss fűszereket, citromhéjat, sót, borsot és kacsazsírt krémmé aprítom. Ezzel kenem be a hús belső oldalát – a bevágásokba is belekerül.
- Bőr előkészítése: A bőrt pár csepp citromlével átkenem, 10 percig állni hagyom, majd papírtörlővel tökéletesen szárazra törlöm. Ezután alaposan besózom és 30 percet pihentetem szobahőmérsékleten.
- Hidegfüstölés: Whiskys tölgyfa forgáccsal 30–40 percig füstölöm, kb. 20 °C-os környezetben. A füst ízmélységet és felszíni szilárdságot ad, nem hőkezel.
- Sütés: A húst feltekerem, spárgával megkötöm, a zöldségágyra helyezem, felöntöm vízzel és borral, majd lefóliázva sütöm 160 °C-on, amíg a maghő nem éri el a 75 °C-t.
Ezután rácsra helyezem a tekercset és 220–230 °C-on grillezem, amíg a bőr arany és ropogós nem lesz. Nem locsolgatom, nem glazirozom – a száraz hő kell a roppanóssághoz. - Pihentetés és tálalás: Szeletelés előtt 15 percig pihentetem szabadlevegőn. A zöldséges szaft szűrve kitűnő mártásként használható, esetleg sűrítve.
🥔 Köretek
Hidegfüstös karalábépüré
300 g karalábét
100 g Gruyere sajt (kiváltó: 70% Cheddar vagy Edami + 30% Parmezán vagy Grana Padano) igény szerint tejszín
50 g vaj
só, frissen őrölt bors ízlés szerint esetleg szerecsendió (opcionális)
- Füstölés: A nyers, szárazratörölt darabolt karalábét 15–20 percre hidegfüst alá teszem. Ettől enyhe, mély háttérízt kap, ami jól illik a porchettához.
- Párolás: Ezután enyhén sós vízben vagy gőz fölött megpárolom, amíg teljesen megpuhul.
- Pürésítés: Melegen botmixerrel simára dolgozom, majd hozzáadom a sajtot/tejszínt és a (barnított) vajat. Sóval, borssal, szerecsendióval ízesítem.
- Állagbeállítás: Akkor jó, ha krémes, nem folyós. Ha sűrű, pár csepp tejszínnel lazítható. Tálalásig melegen tartom.
Rozmaringos, sous vide–ált burgonya salottával
Steak burgonya:
300 g apró burgonya
2–3 ág friss rozmaring
2 gerezd fokhagyma
50 g vaj
só, bors ízlés szerint
Marinád:
100 g salottahagyma
1 ek vörösborecet
3 ek méz
só ízlés szerint
- Szuvidálás: A burgonyát megmosom, szeletelem 3 cm vastag szeletekre, vákuumzacskóba rtegezem vajjal, rozmaringgal, egy gerezd fokhagymával és kevés sóval. 90 °C-on, kb. 60 percig szuvidálom.
- Pirítás: A vajat serpenyőben felolvasztom, aranybarnára pirítom. Végül friss rozmaringgal és kevés borssal ízesítem.
- Marinált hagyma: A hagymát tisztítás után hosszában félbevágom, leveleire szedem. A marinád hozzávalóit összekeverem. A hagymát sous vide zacskóba tesszem, ráöntöm a marinádot, levákuumozom. Mikrosütőben max. fokozaton 30 másodpercig melegítem.
🍽️ Tálalás
A karalábé pürét tesszük alulra a tányérra, arra helyezzük a ropogós burgonyákat. A burgonyákra pakoljuk a Porchetta szeletet, tetejére pedig a hagymaleveleket.
Jó tudni
– Ha tároljuk, akkor lehűtjük 3 szakaszban: 15 perc szobahőmérsékleten, 15 perc csap hideg vízben, 15 perc jeges vízben. Így szaftosabb marad az étel. Tárolás és felbontás nélkül, a hűtő leghidegebb részén (2-4 C° fok) 28 napig biztonságos. Fagyasztást kerüljük mert az étel minősége nagymértékben romolhat a jégkristályok miatt. Tálalás előtt hidegen tárolt élelmiszert felbontás nélkül 60 fokos víz fürdőben fel kell melegíteni, ezután sütjük készre a receptben leírtak szerint.
– A hidegfüst a felszíni fehérjék részleges koagulációját idézi elő, így a hús és a karalábé egyaránt nedvességmegtartóbb lesz. A citromlé enyhe savas közege bontja a kollagént és elősegíti a kérgesedést. A kacsazsír stabilabb hővezető, mint az olívaolaj, ezért a belső rostok lassabban, egyenletesebben főnek át. A száraz sütőtér és a hőgradiens a ropogós bőr záloga – a fizikai–kémiai hatások itt szó szerint hallhatóvá válnak.
 Follow
Follow
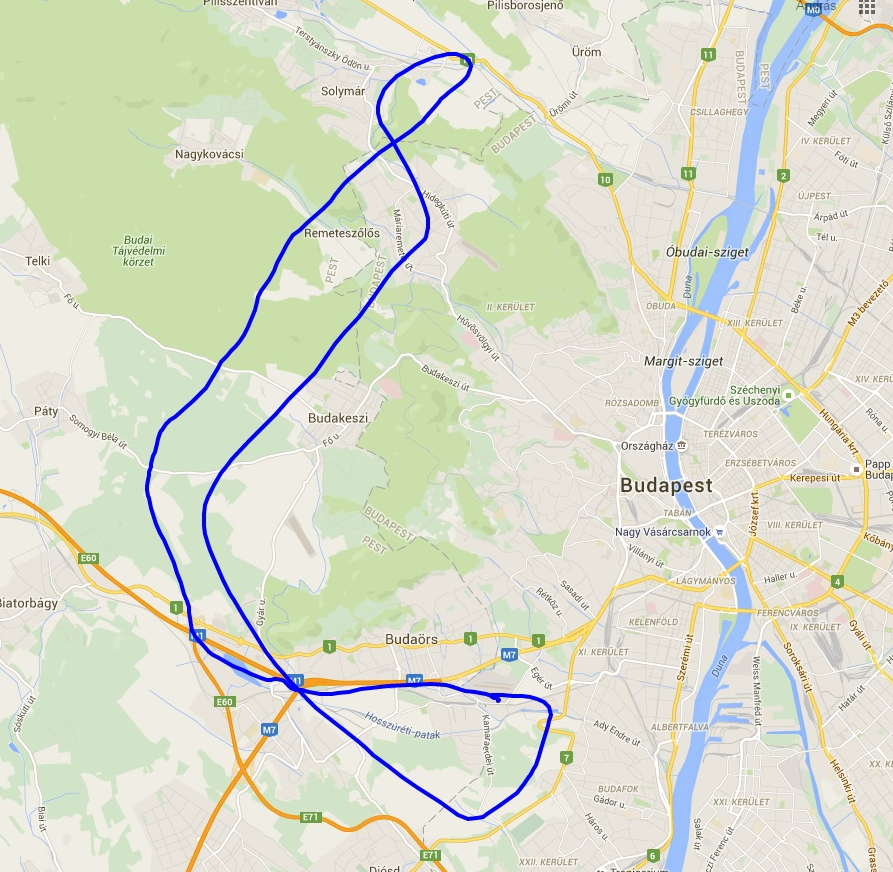
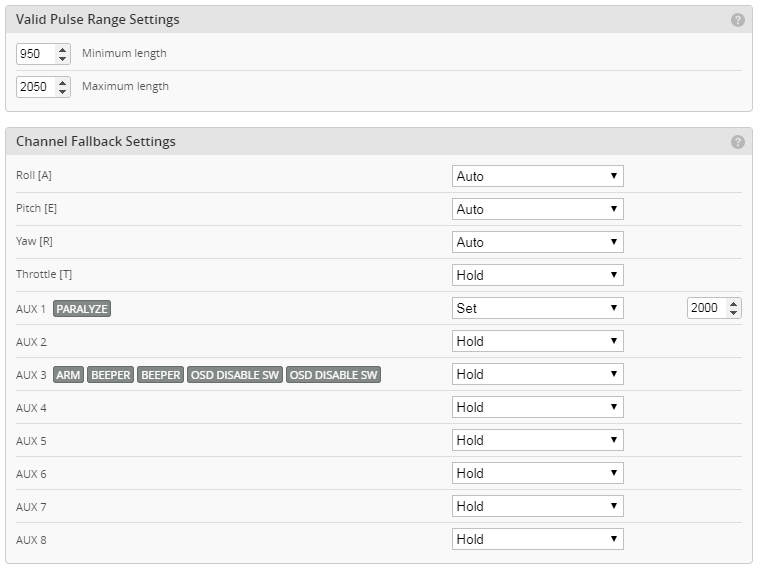
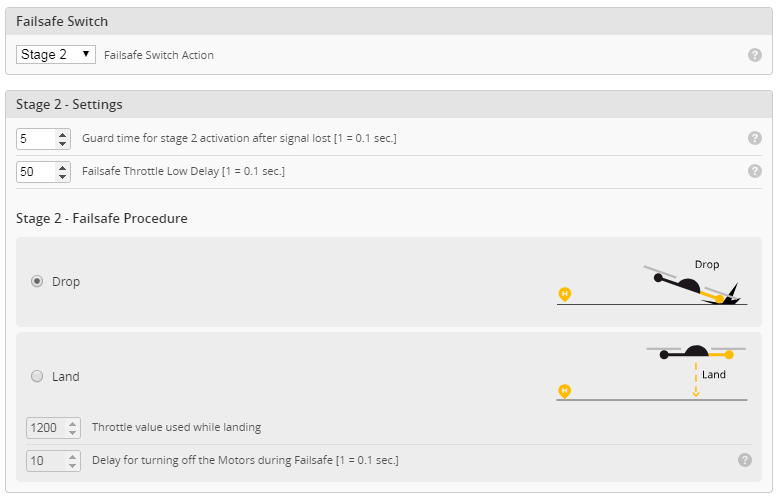
 Az alkatrészeket egy fadobozban kapjuk, ami nemcsak védi, de az előre perforált lemezekből később egy tartót tudunk összerakni, hogy a gépet esetleg ne a párás fűbe kelljen lerakni és a súlypont beállítását is segíti. A teljes kit tartalmazza fa alkatrészeket, szén farokcsövet, tolórudakat és szerelékeket, valamint egy szilikonos felületű nyomtatott építősablont. Ezen nagyon egyszerű a ragasztás.
Az alkatrészeket egy fadobozban kapjuk, ami nemcsak védi, de az előre perforált lemezekből később egy tartót tudunk összerakni, hogy a gépet esetleg ne a párás fűbe kelljen lerakni és a súlypont beállítását is segíti. A teljes kit tartalmazza fa alkatrészeket, szén farokcsövet, tolórudakat és szerelékeket, valamint egy szilikonos felületű nyomtatott építősablont. Ezen nagyon egyszerű a ragasztás. Az építést a szárnyal kezdtem. Az S4083 egy enyhén ívelt profil, ezért a pontosabb építés miatt a bordákat fejjel lefelé rögzítjük az építősablonra. A kis lábaikat egyszerűen a megfelelő téglalapokra ragasztjuk egy csepp pillanatragasztóval és egészen addig így dolgozunk, amíg a szárny alsó torziója felkerül. A bordák függőleges rögzítését egy rétegelt sablon is segíti.
Az építést a szárnyal kezdtem. Az S4083 egy enyhén ívelt profil, ezért a pontosabb építés miatt a bordákat fejjel lefelé rögzítjük az építősablonra. A kis lábaikat egyszerűen a megfelelő téglalapokra ragasztjuk egy csepp pillanatragasztóval és egészen addig így dolgozunk, amíg a szárny alsó torziója felkerül. A bordák függőleges rögzítését egy rétegelt sablon is segíti.